养鱼,本质上是在养水;而养水,往往意味着高昂的电费、人力与技术成本。 在现代集约化与高密度水产养殖中,能耗正悄无声息地吞噬着宝贵的利润。多项行业运营审计显示,在不少现代养鱼项目中,能源成本占总运营支出(OPEX)的比例竟然高达 50%,而其中大头全花在了曝气与增氧系统上。对于中大型养殖场来说,每年几万甚至几十万美元的刚性电费,直接榨干了本可以用来升级饲料或数字化设备的流动资金。 不同养殖模式的电耗差距极大。设计优良的高效系统中,每生产一公斤鱼的电耗可以控制在 $1.0\sim1.8\text{ kWh/kg}$ (Hoseini et al., 2021)。但在一些过度依赖高频循环和复杂机械过滤的工厂化循环水(RAS)中,这个数字在实际运行中甚至会飙升到惊人的 $20.4\text{ kWh/kg}$ (Hoseini et al., 2021)。即便是靠原位微生物自净、号称低过滤需求的生物絮团系统(Biofloc),平均电耗也维持在 $7.2\text{ kWh/kg}$ 的高位 (Hoseini et al., 2021)。在这些复杂的系统架构里,曝气增氧通常占总能耗的 9% 到 37% (Hoseini et al., 2021)。这些数字背后,是真实的白银流向,曝气增氧系统显然已经成了现代养殖业挥之不去的“能耗黑洞”。 我们在科研和一线工程实践中,经常听到一句盲目的经验谈:“溶氧(DO)越高越安全,曝气机开得足存活率才会高。” 这种对超饱和高氧水体的片面追求,不仅违背了物理传质的基本规律,更造成了巨额的电能浪费。从环境工程的第一性原理来看,水体中的溶解氧绝非一个孤立的物理指标,而是连接物理设备传质、水生动物生理代谢与微生物水质净化这三大系统的核心纽带。无节制地曝气,非但无法显著提升接近饱和的水体溶氧,反而会破坏水体微生态,甚至给鱼虾造成物理损伤和严重的应激反应。粗放的运行模式,正在悄悄削弱养殖场的盈利能力,破坏溶解氧系统的稳定性,从而成了限制放放养密度进一步提升的瓶颈。 我们来算一笔具体的经济账。 假设有一个面积为 10 公顷、目标产量为 300 吨的现代化高密度对虾养殖场。为了维持最基础的水质与生物自净,系统在运行高峰期需要约 300 kW 的连续曝气支持 (Sprintex, 2024)。在 150 天的单一养殖周期内,该曝气系统将累计消耗高达 1,080,000 kWh 的电能 (Sprintex, 2024)。如果按新加坡等电力成本较高地区约 0.30 美元/kWh 的标准电价计算,仅曝气一项的电费支出就高达 324,000 美元 (Sprintex, 2024)! 32.4万美元,这绝非一个小数目。 行业数据也印证了这一常态。比如澳大利亚对虾养殖场,其平均曝气能耗长期维持在每吨产品消耗约 4 MWh 电能的水平 (Sprintex, 2024)。然而,出于对缺氧排塘、藻类暴死(“倒藻”)等突发灾难的恐惧,许多养殖户习惯把溶解氧长期维持在 $8.0\sim10.0\text{ mg/L}$ 的近饱和或超饱和状态。这种“宁多勿缺”的盲目心理,不仅让曝气设备成了吞电怪兽,也彻底阻碍了系统整体效率的提升。 更深层次的博弈,在于溶解氧浓度直接支配了水体氮循环的效率。 以自养硝化反应为例,当水体溶解氧浓度在 $3\text{ mg/L}$ 以上时,硝化效率能够逐步提升至 95% 以上,此时氨氧化反应显著增强;学术界关于极限溶氧的研究也指出,硝化作用速率会随着 DO 浓度的升高而增加,直至达到 $3.0\sim4.0\text{ mg/L}$ 之间的阈值,系统才能释放出最大硝化速率 (Zhang et al., 2021)。 但如果溶氧降到 $1.0\sim2.0\text{ mg/L}$ 的低水平,氨氧化与亚硝酸盐氧化过程都会大打折扣,导致硝化反应不完全,并在出水中产生游离氨(Free Ammonia, $NH_3$)。当系统中的游离氨(以 $NH_3\text{-N}$ 计)浓度维持在 $1.0\sim3.0\text{ mg/L}$ 时,会对亚硝酸盐氧化菌(NOB)产生选择性抑制,从而造成毒性极强的亚硝酸盐($NO_2^-$)在水体中大量累积,严重威胁养殖生物的安全。 相反,如果把溶氧一味推高到 $6.0\text{ mg/L}$,好氧反硝化速率又会受到显著抑制,导致总氮(TN)无法有效转化为氮气($N_2$)释放,进而在系统内累积高浓度的硝酸盐 (Zhang et al., 2021)。 因此,在同步硝化反硝化(SND)生物技术中,溶解氧的调控是一门精妙的“平衡艺术”。 大量的实证数据表明,SND 菌群在缺氧和好氧环境中都能稳定存活。但具体到反应速率上,溶解氧高于 $3\text{ mg/L}$ 时,菌群表现出更高的直接氨氧化效率;而在溶解氧低于 $4\text{ mg/L}$ 的区间时,好氧反硝化速率则会显著升高;一旦溶解氧高于 $6\text{ mg/L}$,由于高浓度氧对反硝化酶活性的生物抑制作用,好氧反硝化速率就会受到明显的阻隔。 怎么在单段反应器中平衡这个矛盾? 研究表明,在连续曝气生物膜反应器中,SND 过程的最佳 DO 浓度计算值约为 $3.0\text{ mg/L}$(非常接近实际运行中的 $3.2\text{ mg/L}$)。在此溶氧水平下,系统可实现高达 88.0% 的总氮去除率和 92.3% 的总无机氮(TIN)去除率。 这为我们提供了一个关键的科学启示:通过对溶解氧的第一性原理剖析,将水体溶氧锁定在兼顾鱼虾生理需求、SND生物脱氮黄金窗口与物理传质极限能效的“最优区间”(例如 $2.0\sim3.5\text{ mg/L}$,或特定高负荷系统中的 $3.0\sim4.0\text{ mg/L}$),才是水产降本增效与低碳转型的核心突破口。 * * * ## 溶解氧的物理化学第一性原理与热力学平衡 溶解氧在水体中的行为,其物理化学本质受控于热力学平衡与界面传质的基本物理定律。气体分子向液相的溶解首先遵循亨利定律,即在恒温状态下,某种气体在一定体积液体中的平衡溶解浓度与该气体在液相界面的平衡分压成正比 。在物理传质过程中,吸收系数($\alpha$)是描述特定温度和气压下,单位体积水中溶解气体积的无量纲参数 。下表展示了标准大气压下氧气在水中的吸收系数随温度变化的对应关系 :  在实际水产养殖工程中,为了提高计算精度,通常采用本森-克劳斯(Benson & Krause, 1984)方程,或由加西亚-戈登(Garcia & Gordon, 1992, 1993)修正后的溶解氧饱和度计算模型,该模型全面引入了温度、盐度和气压等多变量纠偏机制 。盐析效应(Salting-out Effect)是影响溶解氧平衡的另一核心机制。高盐度水体中溶解的无机离子(如 $\text{Na}^+$、$\text{Cl}^-$)具有极高的水合能,会强烈吸引极性水分子形成水合离子层,从而压缩非极性的氧气分子在水分子晶格空隙中的生存空间,导致高盐度水体的平衡饱和溶解氧显著低于同等温度下的淡水水体 。此外,大气压力(或海拔高度)通过改变空气中氧气的分压来线性调节饱和溶解氧浓度,静水压则随水深增加而增加,这些因素共同决定了不同空间尺度下的氧气饱和极限 。 在标准大气压($1.000\text{ atm}$)与零盐度($0\text{ ppt}$)条件下,水中溶解氧的热力学极限饱和浓度随温度的变化呈现非线性递减规律 : 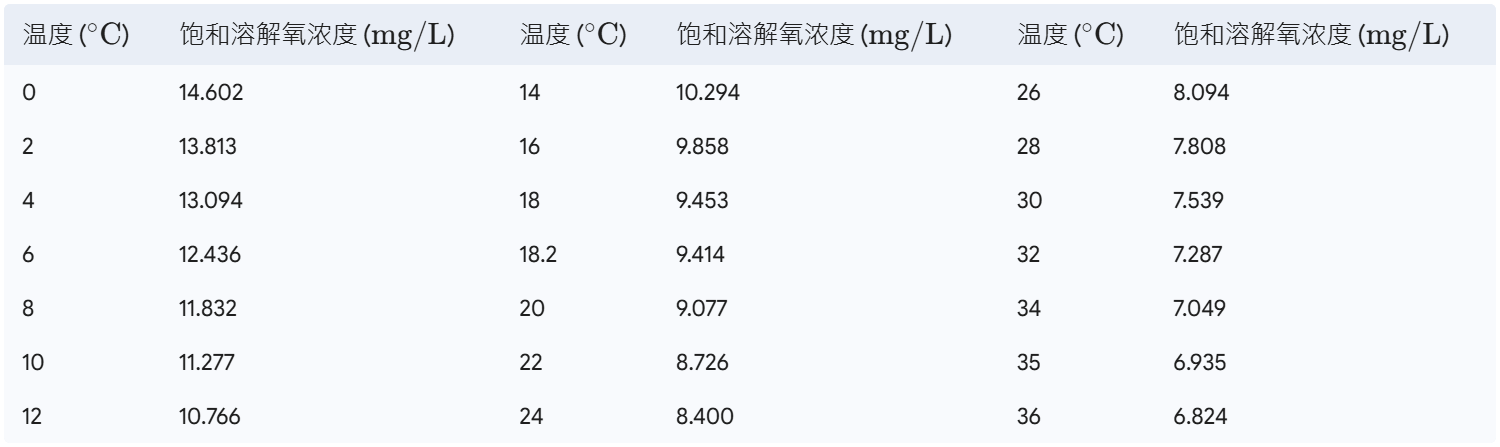 溶解氧是维持水产生物正常有氧呼吸和能量代谢的基础。当环境水体中的溶解氧开始下降时,水产生物会经历一系列从行为调节到急性生理崩溃的过程。评估这些过程的核心生理学指标包括半致死溶解氧浓度($\text{LC}_{50}$)、半亚致死浓度($\text{SLC}_{50}$)以及窒息点 。 传统的将 $2.0\text{ mg/L}$ 定义为水体缺氧临界线的方法存在局限性,超过一半的水生动物在高于此浓度的环境下就已经产生了亚致死或致死生理响应 。 ### 典型养殖品种的生理适宜区间与缺氧表征 在集约化水产养殖中,不同品种的适宜溶解氧范围和缺氧致死点差异显著。如下表所示 : 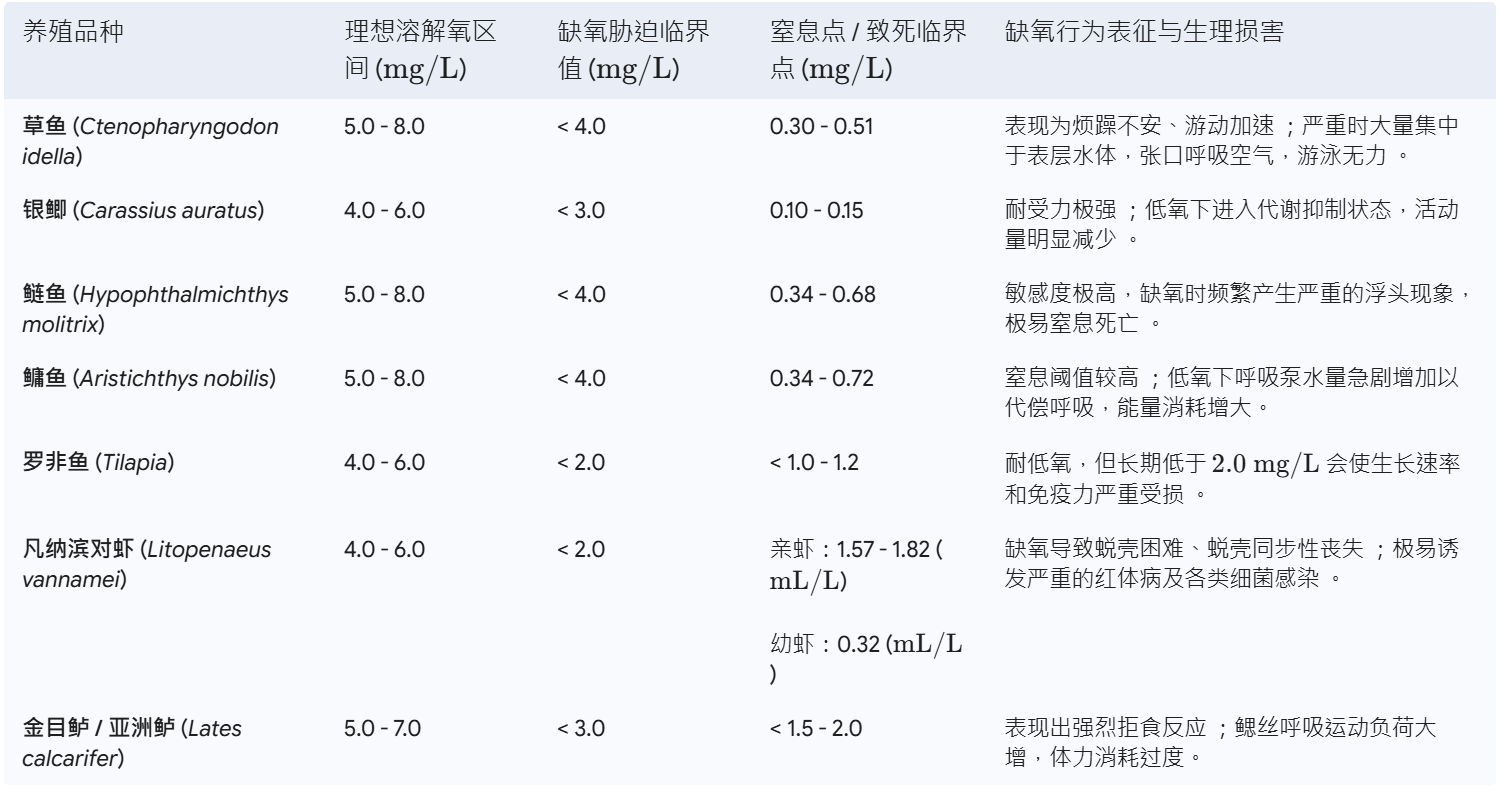 在呼吸调节型鱼类中,当水体溶解氧浓度跌破临界阈值时,机体无法通过单纯提高泵水速率来代偿,开始被迫启动效率地下的无氧糖酵解。由于无氧代谢会产生大量的乳酸等酸性中间体,血液和组织液的酸碱平衡受到破坏,引发代谢性酸中毒。这种酸中毒会引发呼吸系统中的“玻尔效应”和“根效应” 。随着血液中二氧化碳浓度升高、$\text{pH}$ 值下降,血红蛋白与氧气的亲和力以及最大载氧能力大幅降低 。即使此时水体中仍有微量氧气,血红蛋白也无法有效结合并运输氧气,从而在微观层面上加速了细胞缺氧坏死的过程。 同时,慢性缺氧会导致水产动物体内促肾上腺皮质激素释放因子大量分泌,皮质醇水平激增。高水平的皮质醇会强烈抑制免疫细胞的正常增殖,显著降低超氧化物歧化酶(SOD)、溶菌酶等非特异性免疫酶的活性,降低机体对弧菌、爱德华氏菌等病原体的抵御能力,使养殖生物在低氧逆境后的发病率和死亡率呈指数级上升 。 ## 物理化学视角的氧传质阻力与能耗本质 要从根本上破解曝气的“能耗黑洞”,我们必须首先回到传质动力学的第一性原理,剖析氧分子在气-液两相界面转移的底层物理规律与能量耗散机制。 在水处理工程中,氧气向水体的转移过程可以用经典的气液双膜理论来解析。根据该理论,在气液两相界面上,分别存在着高度滞流的气膜(Gas Film)和液膜(Liquid Film)。氧分子必须通过分子扩散依次克服这两层膜的阻力,才能进入主体水体。由于氧气属于难溶气体,其气膜传质系数远大于液膜传质系数,因此气液界面的传质阻力几乎完全集中在液膜一侧。 水体能够溶解氧气的上限,即饱和溶解氧浓度($C_s$),严格遵循**亨利定律(Henry's Law)**: $$C_s = H \cdot P_{O_2}$$ 其中,$H$ 为倾斜常数(亨利常数),$P_{O_2}$ 为气相中的氧分压。$C_s$ 的数值受温度、盐度及大气压的非线性耦合影响。在标准大气压下,随着水温或盐度的升高,$C_s$ 将呈现显著的非线性下降。例如,纯水在 $20^\circ\text{C}$ 时的饱和溶氧约为 $9.08\text{ mg/L}$,而当水温升至 $30^\circ\text{C}$ 时,该数值会降至约 $7.53\text{ mg/L}$;如果是在盐度为 $30\text{ ppt}$ 的海水中,这一上限将被进一步压缩。 这意味在高温或高盐度的集约化养殖环境下,物理传质的天然瓶颈被进一步收窄,系统容错率显著降低。 在实际工程设计中,衡量曝气系统传质能力的核心指标是**氧传质速率(Oxygen Transfer Rate, OTR)**。其动力学基本公式为: $$\text{OTR} = K_La \cdot (C_s - C_L)$$ 式中,$K_La$ 为总传质系数($\text{h}^{-1}$),它反映了曝气设备的机械剪切能力、气液接触比表面积及液膜更新速率;$C_s$ 为饱和溶解氧浓度;$C_L$ 为水体实际溶解氧浓度。 由公式可知,氧传质的根本推动力源于两相界面的浓度差(即溶解氧亏损值,$\Delta C = C_s - C_L$)。