在全球人口爆炸性增长与自然捕捞渔业资源日益枯竭的宏观背景下,水产养殖业已成为支撑全球人类高质量蛋白质供应的最核心支柱。数据表明,在过去的数十年中,水产养殖作为增长最快的食品生产部门之一,其年均增长率一度逼近10%,远超传统畜牧业的3%和捕捞渔业的1.6% 。然而,伴随高密度集约化养殖模式的普及,不可调和的生态环境压力与水质恶化问题日益凸显。其中,养殖水体内部的氮素污染(主要表现为剧毒的氨氮和亚硝酸盐的持续累积)不仅是制约养殖生物产能的物理化学瓶颈,更是诱发大规模病害、引发周边水体富营养化以及威胁人类食品安全的始作俑者 。 为从根本上化解这一系统性矛盾,全球水产养殖水处理系统市场,尤其是循环水养殖系统(RAS)正经历前所未有的爆发式增长。在传统开放式池塘养殖模式下,应对水质恶化的唯一手段是高频次的大量换水,这不仅消耗极其宝贵的淡水或近海清洁水资源,还不可避免地将富含未消化饵料、排泄物、化学药剂和抗生素残留的尾水直接排入自然生态系统 。而现代循环水养殖系统(RAS)通过高度集成的内部水处理机制,能够将换水量削减90%至99%,实现水资源的最大化闭环利用 。 从市场规模的量化数据来看,全球水产养殖水处理系统及循环水养殖系统(RAS)市场在2024年的估值已达到70.5亿美元,预计到2025年将攀升至77.34亿美元,并以8.9%的复合年增长率(CAGR)在2034年突破143亿美元大关 。更有行业分析机构在更为宽泛的统计口径下预测,2025年该市场规模将达到216.3亿美元,至2032年将飙升至484.2亿美元,复合年增长率高达12.2% 。这一庞大且高速增长的市场由多家跨国巨头共同主导,如AKVA group、Pentair PLC、Veolia Group、Water Management Technologies等 ,市场呈现出高度集中化与技术密集型的双重特征 。同时,包括pH、溶解氧(DO)、氨氮、亚硝酸盐在内的水质检测仪器与试剂市场(如检测试剂盒、数字仪表、传感器等)也随之迎来了广阔的增量空间 。 在这样庞大的资本和技术驱动下,目前市场上涌现出了极其繁杂的氨氮和亚硝酸盐去除产品,涵盖了从简单的物理矿物吸附剂、强效化学氧化剂到复杂的基因工程微生物制剂及自动化生物滤池。然而,并非所有技术都能在保证经济效益的同时兼顾长期的生态环境保护与严苛的食品安全合规要求。本报告旨在从物理、化学和生物的第一性原理出发,解构这些市场主流去氮技术的微观作用机理,并基于生态环境保护的韧性要求以及最终走向人类餐桌的食品安全标准,对各类技术路线进行极其深度的剖析与优劣势评估。最终,本报告将引入具有代表性的前沿渔业水处理科技企业——新加坡WaterDoctor,对其基于人工智能(AI)驱动的同步硝化反硝化(SND)生物技术进行全面的商业与技术价值解析。 ## 一、 水产养殖氮素污染的致病机理与第一性原理剖析 ### 1.1 氮素循环的底层机制与能量代谢转换 氮循环是维持全球生物地球化学平衡的基石。在大自然中,氮气($N_2$)通过固氮作用转化为氨($NH_3$),随后参与到复杂的生物代谢网络中 。在水产养殖的具体微观环境中,氮的输入主要来源于高蛋白的人工配合饲料。然而,受限于鱼类和甲壳类动物的肠道消化吸收效率,养殖投入的蛋白质饲料中,约有高达75%的氮素营养无法转化为可收获的动物组织生物量 。这部分庞大的氮素最终以未被食用的残饵、粪便固体颗粒以及直接通过鳃部排泄的溶解态代谢物形式,持续不断地向水体中倾泻 。 在水体底泥或悬浮物中,异养型微生物通过氨化作用,将这些复杂的含氮有机大分子(如蛋白质、氨基酸)降解剥离,最终释放出总氨氮 。这是整个水产养殖氮素危机的源头。总氨氮(TAN)在水溶液中并非以单一形态存在,而是基于酸碱离解常数,在剧毒的非离子氨($NH_3$)与相对无毒的离子铵($NH_4^+$)之间维持着动态的化学平衡 。 其第一性原理的化学平衡方程式表述为: $NH_3 + H_2O \rightleftharpoons NH_4^+ + OH^-$ 这一化学平衡不仅是一个静态过程,更是极度敏感于水体环境变量的热力学动态过程。温度和pH值的升高会强烈驱动反应向左进行,导致水体中剧毒的非离子氨($NH_3$)比例呈指数级上升 。非离子氨的分子结构使其不带有电荷且具备高度脂溶性,这使得它能够毫无阻碍地穿透水生动物鳃部的脂质双分子层细胞膜,直接进入动物血液循环系统 。在血液中,高浓度的氨会干扰中枢神经系统,破坏细胞内的渗透压平衡,引发严重的能量代谢紊乱,不仅抑制动物生长,长期暴露更会引发大规模死亡 。科学研究表明,大多数水产养殖系统中非离子氨的可接受安全阈值极低,仅为0.025 mg N/L 。此外,氨的积累还会在底泥沉积物中产生显著的含氮生化需氧量,因为微生物在尝试氧化这些氨氮时,会疯狂消耗水体中的溶解氧,导致水体急剧缺氧甚至引发翻塘事故 。 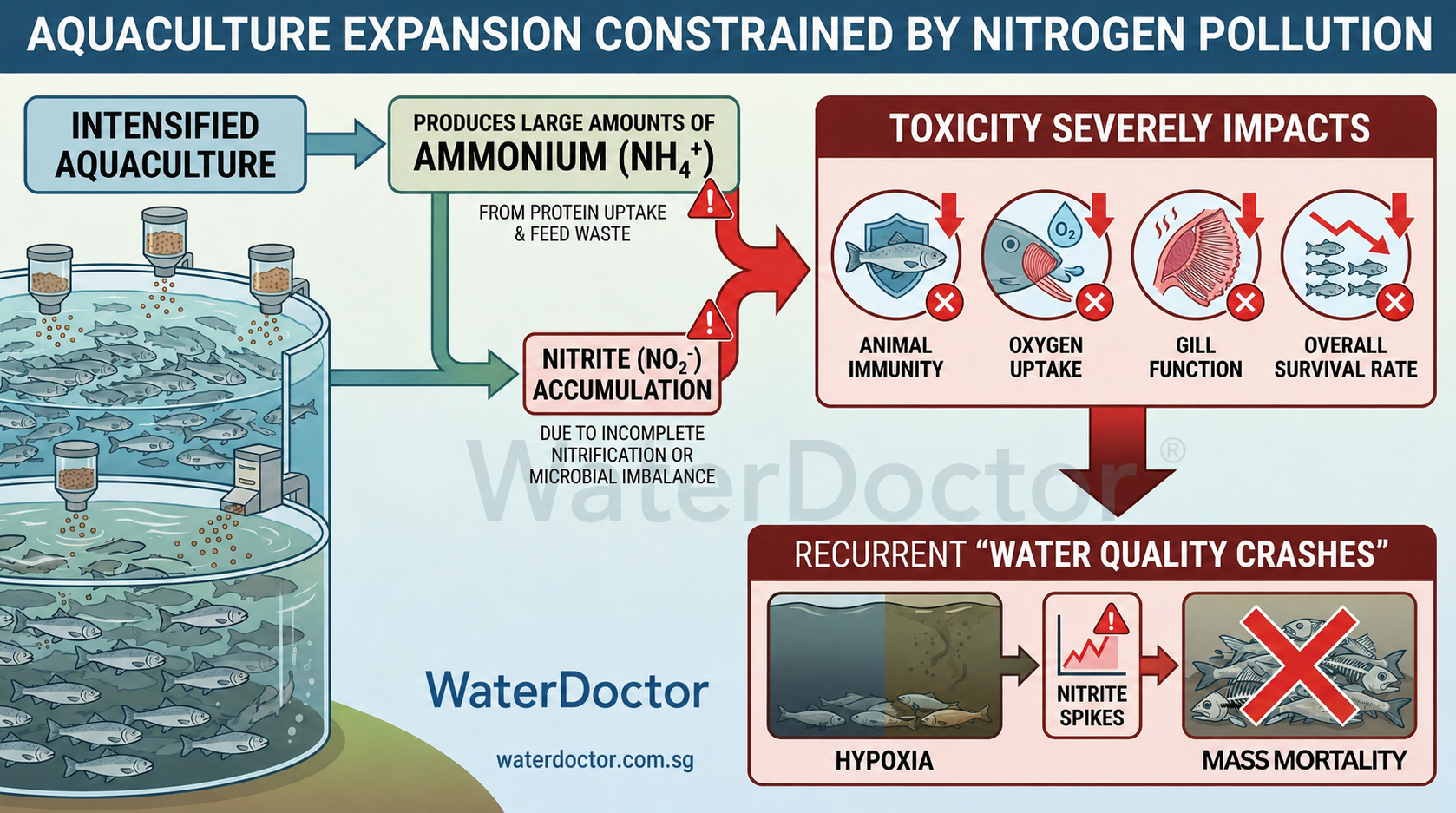 ### 1.2 亚硝酸盐的分子毒性与酶学氧化机制 当水体中的氨氮试图通过自然界的硝化作用(Nitrification)进行自我解毒时,氮素循环进入了第二阶段。水体中自然存在的氨氧化细菌(Ammonia-oxidizing bacteria, AOB,代表菌属为Nitrosomonas)利用溶解氧,将氨或铵离子初步氧化为亚硝酸盐($NO_2^-$) 。在正常的生态位中,亚硝酸盐氧化细菌(Nitrite-oxidizing bacteria, NOB,代表菌属为Nitrobacter和Nitrospira)会迅速接棒,将亚硝酸盐进一步氧化为相对无毒的硝酸盐($NO_3^-$) 。 然而,这种串联的生物化学反应在集约化水产养殖中常常出现“脱节”或“反应速率不匹配”,导致中间产物亚硝酸盐在水体中危险蓄积 。这主要是由于AOB和NOB在世代繁殖时间、对底层环境的敏感度上存在差异。例如,Nitrospira等NOB的世代生长周期长达9.87小时,其指数生长期内的亚硝酸盐氧化速率仅为4.63 pg $NO_2^-$-N/细胞/小时,且对溶解氧浓度(当DO接近1.0 mg/L时完全受到抑制)、盐度波动甚至特定波长的光照(如410 nm至520 nm波长会引发光抑制现象)具有极高的敏感性 。 亚硝酸盐一旦积累,其对鱼类等水生动物的分子毒理机制将极其致命。亚硝酸盐通过鱼鳃被主动吸收到血液后,会作为一种强氧化剂,直接与红细胞中的血红蛋白(Hemoglobin)发生不可逆的氧化还原反应。它将血红蛋白中心原卟啉环上的二价铁离子($Fe^{2+}$)氧化为失去携氧能力的三价铁离子($Fe^{3+}$),形成高铁血红蛋白(Methemoglobin) 。这一病理过程在水产医学上被称为“褐血病”(Brown Blood Disease),受感染的鱼类会表现出严重的组织缺氧、窒息浮头,最终因多器官衰竭而死亡 。 值得注意的是,从分子酶学的第一性原理来看,负责催化亚硝酸盐氧化为硝酸盐的亚硝酸盐氧化还原酶(NXRs)与将硝酸盐还原的硝酸盐还原酶(NARs)在酶学结构和氧化还原机制上具有惊人的相似性,两者均依赖于钼(Molybdenum)辅酶基团,且氧原子转移(Oxygen atom transfer, OAT)是其共同的核心机制通道 。这种酶学层面的“一体两面”不仅揭示了微生物氮素代谢的底层灵活性,也为后续人工通过调控氧化还原电位或环境条件,实现特定微生物除氮提供了坚实的生化理论基础 。 ## 二、 市场主流氨氮/亚硝酸盐去除技术的机理解析与应用格局 面对上述严峻的理化毒性挑战,水产养殖从业者和科技企业开发了多条技术路线。根据干预介质的物理化学属性,目前市场上的去除技术主要可以划分为三大阵营:物理吸附截留法、化学氧化还原法以及传统及现代生物转化法。 ### 2.1 物理吸附法:离子交换与晶体分子筛的结构力学 以天然矿物沸石(Zeolite,特别是斜发沸石 Clinoptilolite)为绝对代表的物理法,是历史最悠久且应用广泛的水质急救方案之一 。 沸石之所以能够去除水中的氨氮,完全取决于其独特的硅铝酸盐多孔晶体结构。在晶格内部,由于部分四价硅离子被三价铝离子同晶置换,使得整个晶体骨架带有负电荷。为了保持电中性,晶格孔道内吸附了大量易于游离的碱金属阳离子(如$Na^+$、$K^+$等) 。当富含铵离子($NH_4^+$)的养殖水体流经沸石颗粒时,基于离子交换平衡原理,$NH_4^+$因其特定的水合离子半径与沸石孔径高度匹配,被强力吸附进晶格内部,同时置换出$Na^+$进入水体。其核心可逆反应方程式表述为: $NH_4^+ + R^- Na^+ \Leftrightarrow Na^+ + R^- NH_4^+$(其中$R^-$代表带负电荷的沸石硅铝骨架) 。 除了直接的氨氮吸附,物理结构上的微孔截留还能显著降低水体浊度,吸附硫化氢等引发恶臭的气体,并在一定程度上降低生物絮团系统(Biofloc)中的污泥沉积 。在一些淡水养殖实验中,向水体中添加15g/L至20g/L的斜发沸石,能够有效遏制氨氮峰值,甚至通过其离子交换作用削弱某些重金属(如镉)在鱼类组织中的生物富集 。 **生态与物理局限性**:从第一性原理来看,物理吸附存在不可逾越的热力学与空间位阻(Steric hindrance)障碍。首先是竞争性吸附问题。在海水、半咸水或硬水养殖环境中,水体中天然存在着极高浓度的钠离子($Na^+$)、钙离子($Ca^{2+}$)和镁离子($Mg^{2+}$)。这些阳离子会与$NH_4^+$展开激烈的吸附位点争夺,导致沸石的除氨效率呈现断崖式下跌,这使其在海洋循环水养殖或海水池塘中基本失效 。其次,物理方法仅仅是污染物的“空间转移”而非“物质消解”。一旦沸石的离子交换容量达到饱和,它不仅失去净化功能,若将其作为底泥长期留在池塘中,当水体理化条件(如盐度或pH)发生突变时,沸石可能发生逆向解吸,瞬间向水体释放大量此前囤积的氨氮,构成严重的生态定时炸弹 。 ### 2.2 化学氧化法:强氧化还原反应与自由基DNA破坏 化学氧化剂因其起效快、兼具广谱杀菌作用,在传统的土塘养殖和病害爆发期备受养殖户青睐。当前市场上的主流化学除氮剂包括单过硫酸氢钾复合盐(Potassium Monopersulfate, 简称KMPS或MPS)、二氧化氯($ClO_2$)以及过氧化氢($H_2O_2$)复合物 。 以单过硫酸氢钾复合盐(KMPS)为例,这是一种非氯强氧化剂,其在水中溶解后能够迅速释放出新生态的活性氧自由基以及高电位的氧化基团 。它的除氨氮和亚硝酸盐机制分为直接与间接两条路径。间接方面,KMPS能够强力氧化并分解积聚在池塘底部的残饵、粪便等大分子有机质,从源头上削减了氨氮生成的前体物质来源 。直接方面,KMPS产生的强氧化自由基能够直接穿透微生物的细胞壁,与细胞内遗传物质发生不可逆的氧化还原反应,导致细菌的DNA和RNA结构发生变性与突变,从而彻底摧毁微生物的繁殖和生长能力 。另一条路径则是基于过氧化氢($H_2O_2$)等制剂,在搅拌作用下缓慢释放氧气,在局部高氧化电位下,通过纯化学途径直接将亚硝酸盐氧化为相对无毒的硝酸盐,进而被后续投放的光合细菌(PSB)作为营养物质吸收 。同时,二氧化氯($ClO_2$)在0.02至0.5 ppm的极低浓度下,也被证实可以高效降低细菌负荷并控制有害藻华,保持水质清澈 。 **生态灾难与毒理学反噬**:从生化反应的非特异性特征来看,化学氧化剂是一把极度危险的“双刃剑”。由于化学反应缺乏生物酶那种精确的靶向识别能力,其强氧化性表现出彻底的“无差别杀伤”。大量的微观生态测序研究揭示了滥用KMPS的严重后果。在添加了3 mg/L至8.